
別項Ⅱ ニホンザル・群れ識別法による長期調査から見えてきたこと
これは本年(2026年)1月に京都市で開催された「霊長類学史研究会」での講演の要旨で、日本のサル学黎明期から続けられてきた“個体識別法”でなく、“群れ識別法”による長期調査でなにが明らかになったかについてである。
調査地と初期の研究
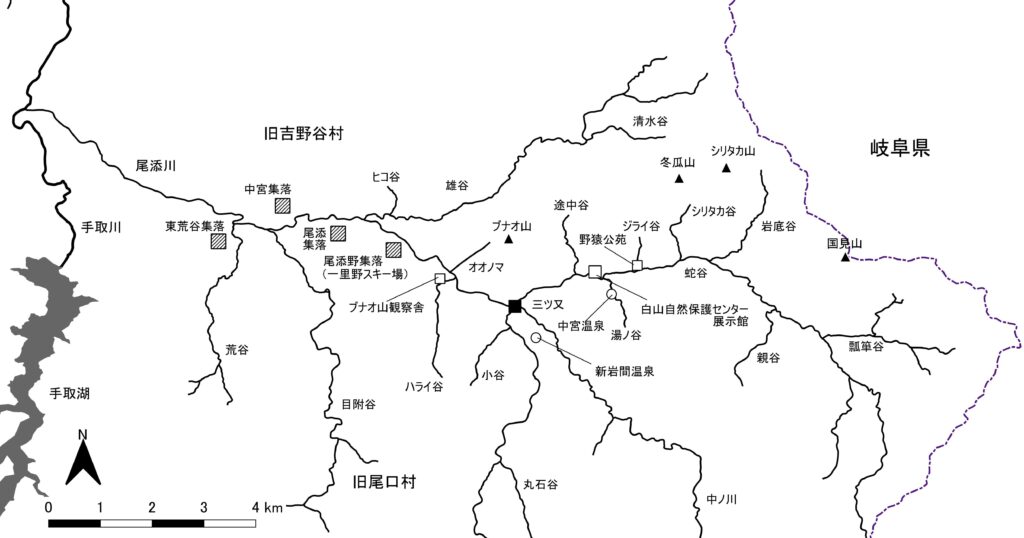
白山北部山域の石川県側、手取川右岸の支流尾添川流域(図1)に生息するニホンザルの組織立った研究は、1964年から河合雅雄、川村俊蔵らによって開始され、1966年からは河合をリーダーとして日本モンキーセンター(以下、JMC)の研究員や学芸員によって鋭意進められてきた。筆者は東アフリカでのチンパンジー調査から1967年6月に帰国し翌年JMCに就職。その年(1968年)の秋からJMC調査団に加わった。


群れはいくついるか
1964年からの調査成果は、筆者が参加した当時すでにおおよそがまとめられていて、群れ分布に関しては尾添川流域に8群生息すると結論付けられていた(『白山の自然』 末尾参照)。8群のうち2群は1966年に地元の人によって餌付けされた群れ(「カムリA群」)と、餌付け途中に分裂し消息を絶った群れ(「カムリB群」)である。残り6群はいずれも積雪期には山奥の標高の高い所から谷や沢の低い所に下りて来るという。
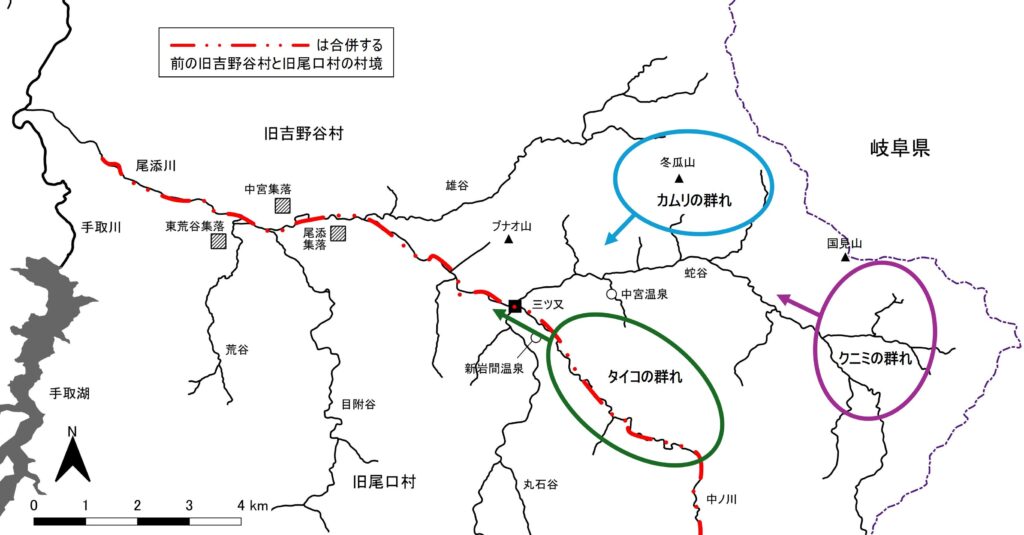
続く1969年2月と翌年2月には一人この流域で調査したが、テーマは群間関係で、複数の群れの同時追跡が必須だから、雪深い急斜面に6群をひたすら探し求め、結局餌付いた「カムリA群」のほか「タイコの群れ」と「クニミの群れ」の2群しかいないことが明らかになった。群れの存在は出会えればそれで済むが、いるといわれた群れが実際には存在しないことを証明するのはじつに大変な作業だった。
群れ識別法
筆者は冬期調査を現在も続けているが、調査方法は日の当たる南斜面に広がる群れを対岸から観察し続けることだ。当然、対象のサルたち1頭1頭は遠すぎて個体識別など無理である。しかし、来る日も来る日も群れ全体を視覚でしっかり捉え続けていると、山ほどの映像を通して、やがて“群れ”が脳裏で概念化されていく。そうするとニホンザルの生きざまの新しい側面が鮮明に浮かび上がってくるものだ。
群れの存在のフェイズ
群れは個体の寿命をはるかに越えて存在する。その存在のフェイズは、頭数が50頭ほどになると群れの広がりが目立って大きくなり、追随オスの数も増える。分派行動も見られ始める。分派集団ははじめは小さく、かつ半日か長くても2日で本体に合流する。もう少し頭数が増えると、大きな分派集団が観察されるようになり、分派集団は遊動域の拡張を始める。分派している期間は1週間とか2週間の長期になることもある。やがて頭数が70~80頭に達すると、頻繁に分派を繰り返していた集団を中心に群れは分裂するが、その前後から遊動域の拡張面積が大きくなる。そして分裂後の遊動域拡大が終わると、二つに別れた群れはそれぞれ落ち着き、最初のフェイズに戻る。ただし、この一連のフェイズは、冬の積雪量や雪崩の落ち方、秋の堅果類の稔り具合など自然現象に左右されるし、隣接群がその時どのフェイズにあるかにも影響される。分裂したどちらかが高飛びすることもときにある。
群れのによる分裂の仕方や行動圏拡大の仕方の違い
筆者はこれまで3群中2群、餌付けされた「カムリA群」と野生群「タイコの群れ」の、分裂の仕方や遊動域拡大の仕方の違いを記録してきた。その違いを図2と図3に示した。図2は1960年代前半の両群の冬から春にかけての遊動域で、頭数は60頭余りでほぼ等しく、遊動域の広さもほぼ同じだった。それが1970年代以降、図3のように両群で一気に変化する。
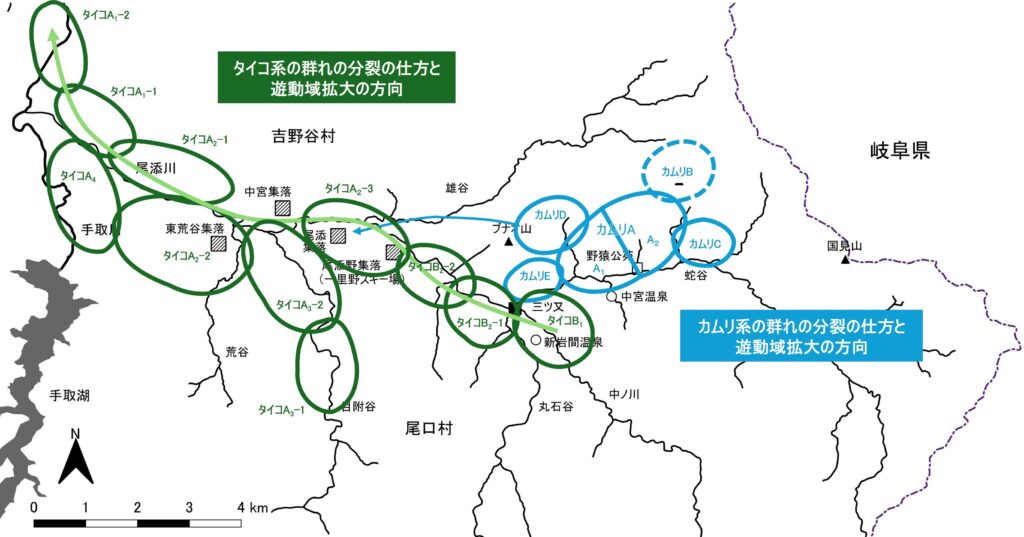
「カムリA群」にとって自然にはない美味で栄養価の高いトウモロコシやダイズが腹一杯食べられる餌場の魅力はなにものにも替え難い。だから分裂群はいずれも小さく、遊動域はいざとなったらいつでも餌場に戻れるよう、主群に対して瘤のようになる(主群と分裂群のすべてを一括して以後“カムリ系の群れ”と呼ぶ)。そして、分裂しても周辺に遊動域を構えるだけの好ましい土地がなくなると、主群は餌場を分け合うように分裂した(図3の「カムリA1群」と「カムリA2群」)。
一方「タイコの群れ」は「カムリA群」が餌付けされたと同じ年から、晩秋に山奥から中ノ川下流域に下りて来ると、新岩間温泉と中宮温泉(図1参照)の冬期閉鎖に合わせてダイコン畑等の取り残しを漁り、余勢をかって三ツ又から往復1日か2日で、ずっと下の尾添集落や中宮集落のカキの実漁りを強行するようになった。そうして頭数が増えて群れが分裂すると、小さい分裂群をこれまでの遊動域に残して主群は下流域へ進出するというパターンを繰り返す(上記“カムリ系”と同様すべてを“タイコ系の群れ”と呼ぶ)。下流へ行けば行くほど集落は大きくなり、カキ等の果実がなっている量も多くなるし、周囲の田畑の面積も桁違いに広くなるからである。
尾添川を挟んでの村人や集落の人々の生活習慣の違い
ここでもう一度、図3を見て欲しい。「カムリA群」の分裂群はのちに高飛びする1群を除き蛇谷流域にとどまって、中ノ川流域や三ツ又より下流までは遊動域を拡大していない。一方「タイコの群れ」は中ノ川から尾添川下流まで遊動域をどんどん広げ、さらに手取川本流まであっという間に到達した。
ところで尾添川を境にして右岸(吉野谷村や中宮集落)と左岸(尾口村や尾添と東荒谷集落)では最近まで、戦国時代以前からの宗教・宗派の違いによる対立や戦の歴史を通して冠婚葬祭をはじめ生活習慣が極端に異なっていた(『白山ー自然と文化ー』 末尾参照)。そうすると、尾添川と蛇谷・中ノ川を境にカムリ系の群れとタイコ系の群れのここまで見てきた分裂の仕方や遊動域拡大の仕方のこれほどまでの違いもまた、ニホンザルを群れという単位で見た時の文化の違いという視点で検討する必要がありそうだ。
集落の内と外というすみわけ
以上述べたこととの関連でもう一例、分裂した当初から注目して追ってきた「カムリD群」を紹介しよう(分裂した1985年当時の頭数は12頭とごく小さい)。
この群れもやはり瘤のように北西方向へ分裂したが(図3参照)、地形がなだらかで冬場の生息環境としては不適だったので三ツ又越えをし、タイコ系3群の遊動域すら越えて(高跳びして)、ブナオ山北西の急斜面(雄谷左岸)に遊動域を構えた。だがそこもまた冬場には雪や風をまともに浴び、めぼしい食物もなかったが、救いは尾添集落や中宮集落に近かったことである。
先に書いたように、タイコ系にとって好ましい生息環境とは、民家がより密集している場所だ。民家の敷地内には漿果や堅果を稔らす木が何本も植えられていて、周囲には採り残しを漁れる田畑が広がっているからである。しかし、警戒する対象の人々も多く住んでいるわけで、採食には人のいない時間帯を選ぶ必要がある。一方カムリ系にとっては餌場が最も好ましい場所で、餌場(野猿公苑)には人がいない時も数人の時も観光客が沢山いる時もあるが、警戒する必要は全くないし、ときには客から餌場に撒かれた以外の美味しいものももらえる。したがってタイコ系が警戒している集落の中に入り込んでしまえば、放置されたカキの実等美味しいものを食べられるし、家から人が出て来ても、とくに冬場なら、近づいて行って少し媚を売るだけでミカンやリンゴの一つが手に入ることもある。
こうして尾添、中宮、東荒谷等の集落の内と外で、タイコ系の複数の群れとは一種の“すみわけ”が成立。集落の中には良好な泊り場も事欠かないから、タイコ系より早いスピードで頭数を増やしていった。すなわち「カムリD群」はタイコ系に囲まれながらも、カムリ系の“習性”を十分発揮して成功を収めた(繁殖率の高さ)ことが理解されよう。一里野スキー場が大変賑わっていた2000年代初め、スキー客が集まるゲレンデ入口のすぐ近くをゆうゆうと採食移動した群れは「カムリD群」だけで記録されているし、来訪者が多いブナオ山観察舎の入口に屯したのもこの群れだけである(当時は50頭ほどの群れになっていた)。
参考文献
『白山の自然』日本自然保護協会中部支部・白山学術調査団編 石川県,1970,pp.395
『白山ー自然と文化』白山総合学術書編集委員会編 北国新聞社,1992,pp.514
(追記)先週の12日(2026年2月)、例年通りの白山のサル冬期調査を開始して4日目の晴れた朝。尾添川下流に遊動域をもつタイコ系の群れの現状を把握するため、一里野スキー場から県道360号線に車を走らせ尾添集落に差し掛かった時のことだ。なんと、カムリD群が集落から出て来て、警戒の素振りも見せずに車のすぐ前を横切っていくではないか。かれらはそのあと、法面の道路脇をのんびりと下流方向へ採食移動していく。
一昨年と昨年と続けて、大型捕獲檻で仲間が何頭も捕獲されたにもかかわらず、今なお人を頼りにする習性を捨てないサルたちに、車窓から移動の様子を目で追いつつ、筆者は束の間の感慨に浸った。
